Phosphatidylinositol 4,5-bisphosphate decreases the concentration of Ca2+, phosphatidylserine and diacylglycerol required for protein kinase C α to reach maximum activity.
Study Design
- Studientyp
- In Vitro
- Population
- Lipid vesicle enzyme assay (in vitro)
- Intervention
- Phosphatidylinositol 4,5-bisphosphate decreases the concentration of Ca2+, phosphatidylserine and diacylglycerol required for protein kinase C α to reach maximum activity. None
- Vergleichsgruppe
- None
- Primärer Endpunkt
- PKC alpha enzyme activation kinetics
- Wirkungsrichtung
- Neutral
- Verzerrungsrisiko
- Unclear
Abstract
The C2 domain of PKCα possesses two different binding sites, one for Ca(2+) and phosphatidylserine and a second one that binds PIP2 with very high affinity. The enzymatic activity of PKCα was studied by activating it with large unilamellar lipid vesicles, varying the concentration of Ca(2+) and the contents of dioleylglycerol (DOG), phosphatidylinositol 4,5-bisphosphate (PIP2) and phosphadidylserine (POPS) in these model membranes. The results showed that PIP2 increased the Vmax of PKCα and, when the PIP2 concentration was 5 mol% of the total lipid in the membrane, the addition of 2 mol% of DOG did not increase the activity. In addition PIP2 decreases K0.5 of Ca(2+) more than 3-fold, that of DOG almost 5-fold and that of POPS by a half. The K0.5 values of PIP2 amounted to only 0.11 µM in the presence of DOG and 0.39 in its absence, which is within the expected physiological range for the inner monolayer of a mammalian plasma membrane. As a consequence, PKCα may be expected to operate near its maximum capacity even in the absence of a cell signal producing diacylglycerol. Nevertheless, we have shown that the presence of DOG may also help, since the K0.5 for PIP2 notably decreases in its presence. Taken together, these results underline the great importance of PIP2 in the activation of PKCα and demonstrate that in its presence, the most important cell signal for triggering the activity of this enzyme is the increase in the concentration of cytoplasmic Ca(2+).
Zusammenfassung
The results showed that PIP2 increased the Vmax of PKCα and demonstrated that in its presence, the most important cell signal for triggering the activity of this enzyme is the increase in the concentration of cytoplasmic Ca2+.
Full Text
Phosphatidylinositol 4,5-Bisphosphate Decreases the Concentration of Ca2+, Phosphatidylserine and Diacylglycerol Required for Protein Kinase C a to Reach Maximum Activity
Antonio L. Egea-Jime´nez, A´ ngel Pe´rez-Lara, Senena Corbala´n-Garcı´a, Juan C. Go´mez-Ferna´ndez*
Departamento de Bioquı´mica y Biologı´a Molecular-A, Facultad de Veterinaria, Regional Campus of International Excellence "Campus Mare Nostrum", Universidad de Murcia, Murcia, Spain
Introduction
PKCa (protein kinase C a) is a classical PKC isoenzyme that is activated by second messengers, namely the increase in Ca2+ concentration in the cytoplasm of the cell and the appearance of diacylglycerol in the membrane, where it establishes specific interactions with phosphatidylserine and PIP2 [1].
The translocation of classical PKCs (cPKCs) to the plasma membrane is mediated by the C1 and C2 domains, and it has been shown that initial membrane affinity is mainly determined by C2 domain–membrane interactions, followed by C1 domain–diacylglycerol interactions [1]. One of the main sources of diacylglycerol in the plasma membrane following cell stimulation is PIP2 which is hydrolyzed by phospholipase C to produce diacylglycerol and inositol 1,4,5-trisphosphate, which together activate protein kinase C for sustained cellular responses [2]. However, it has been shown that PIP2 may also activate PKCa by direct binding to a polylysine motif located in strands b3 and b4 [3–7] and that can be considered a specific site for PIP2 [8] (see Fig. 1). Other molecules like phosphatidylserine or phosphatidic acid [9] or even retinoic acid [10] may also bind with lower affinity to this site. It has been clearly shown that PIP2 is important for PKCa translocation to the
membrane and for prolonging this translocation. Rapid [5,11,12] kinetics studies on the binding of this enzyme to model membranes suggested that the interaction of PKCa with membranes occurs via two steps: a rapid weak recruitment to the membrane due to non-specific interactions with (primarily) anionic lipids and the formation of a high affinity complex due to stereospecific interactions of each PKCa domain with its specific ligands [12].
PKCa enzyme is a paradigmatic example for bearing a C2 domain which may simultaneously bind three different activators, in this case Ca2+, phosphatidylserine and PIP2. Fig. 1 shows this C2 domain in which Ca2+ binds to its site, acting as a bridge for phosphatidylserine, although this phospholipid also directly interacts with several protein residues [13,14]. In another site located in a b-groove, PIP2 binds with great affinity.
Previous work has shown that PKCa exhibits high cooperativity in its activity by phosphatidylserine [15,16] and that the two second messengers of the kinase, diacylglycerol and Ca2+, markedly increase the affinity of the kinase for phosphatidylserine [17]. In this paper, we use highly purified full-length PKCa to perform a kinetic study of the activation of PKCa by model membranes, in which the concentrations of POPS, DOG, PIP2
and Ca2 are varied. Our results indicate that PIP2 enhances PKCa activity and decreases the required concentrations of the other activators, to reach maximum activities.
Materials and Methods Materials
1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphoserine (POPS), 1-palmitoyl-2-oleoyl-sn- glycero-3-phosphocholine (POPC), L-a-phosphatidylinositol 4,5-bisphosphate (PIP2) and 1,2-sn-dioleoylglycerol (DOG) were purchased from Avanti Polar Lipids (Alabaster, ALA, U.S.A.). To discard any 1,2-diacyl-sn-glycerol contamination of the PIP2 source used, L-a-phosphatidylinositol 4,5-bisphosphate (PIP2) (lot BPIP2 (4,5)-54) was analyzed by Avant Polar Lipids and their release testing included Gravimetric analysis, Phosphorus analysis, Proton and Phosphorus NMR, Mass Spectrometry, Thin Layer Chromatography (TLC), HPLC and Calcium by ICP/MS without detecting any impurity. The HPLC assay indicated 100% PIP2. In addition, once dissolved it was also analysed in our laboratory by using TLC and no impurity was detected either. All other chemicals were of high purity and supplied by Sigma Chemical Co. (Madrid, Spain).
Expression and Purification of Protein Kinase Ca
The full length cDNA for rat PKCa was kindly provided by Profs. Ono and Nishizuka (Kobe, Japan). PKCa was cloned into the plasmid pFastBac HT (Invitrogen, Madrid, Spain). A 0.5 litre scale culture of Sf9 insect cells (Spodoptera frugiperda) at 2.16106 cells/ml was infected with the recombinant baculovirus.Cells were harvested 60 h postinfection (cell viability 80%), pelleted at 4500 rpm for 20 min, and resuspended in buffer containing 25 mM Tris-HCl pH 7.5, 100 mM EGTA, 50 mM NaF, 100 mM NaVO3, 1% Triton X-100, 10% glycerol, 150 mM NaCl, 1 mM
PMSF, 10 mg/ml leupeptin and 10 mM benzamidine. The pellet was disrupted by sonication (6610 s) and the resulting lysate was centrifuged at 15000 rpm for 20 min. The supernatant was applied to a 1 ml His-Gravi TrapTMH column (GE Healthcare, Barcelona, Spain) and equilibrated with 25 mM Tris-HCl pH 7.5, 150 mM NaCl and 20 mM imidazole buffer. The bound proteins were eluted by an imidazole gradient (20–500 mM). Fractions containing protein kinase Ca from a His-Gravi TrapTMH column were pooled, concentrated by ultrafiltration to a 2 mL volume and adjusted by the addition of 5 M NaCl to give a NaCl concentration of 1 M.
This fraction was then processed by hydrophobic exchange chromatography, directly applying it to a SOURCE 15PHE 4.6/ 100 PE H columHM Tris-HCl pH 7.5, 1 mM DTT and 10% glycerol. After the unbound proteins had passed through the column, PKC was eluted with a gradient of 0.8–0 M NaCl. Highly pure PKCa was obtained, as determined by SDS-PAGE (12.5%). The protein was aliquoted and stored at 280 C in the presence of 10% (w/v) glycerol and 0.05% (v/v) Triton X-100.
Preparation of Phospholipid Vesicles
Lipid vesicles were generated by mixing chloroform solutions of 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC), 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoserine (POPS) and L-a-phosphatidylinositol-4,5-bisphosphate (PIP2) in the desired proportions. Lipids were dried from the organic solvent under a stream of oxygen-free nitrogen, and then the last traces of organic solvent were removed under vacuum for at least 4 h. Dried phospholipids were resuspended in the corresponding buffers by vigorous vortexing and then large unilamellar phosholipid vesicles of about 100 nm diameter were prepared by extruding (11 times) rehydrated phospholipid suspensions through two stacked 0.1 nm polycarbonate membranes (Millipore Inc., Bedford, MA, USA).
Enzymatic Activity Assay
Enzymatic activity was assayed using a technique described previously [17], in which the incorporation of radioactive phosphate [c-32P] to kinase substrate (histone III-S) was measured. Lipids in organic solutions were mixed in the desired proportions and dried under a stream of N2, removing the last traces of organic solvent by keeping the samples under vacuum for at least four hours. Large unilamellar vesicles (LUVs) were prepared using the extrusion technique, as explained above. These lipids were resuspended immediately before use in a buffer composed of 20 mM Tris-HCl (pH 7.5), 0.5 mg/ml of histone III-S, 40 mM ATP [c-32P] (3000,000 cpm/nmol), 5 mM MgCl2 1 mM EGTA and enough CaCl2 to give a free Ca2+ concentration of 200 mM, except in the assays in which this concentration was varied. The necessary concentrations of CaCl2 added in each case were calculated by using the procedure described in [18]. The final concentration of lipids in the reaction mixture was 625 mM. The reaction was started by the addition of 5 ml of PKCa (0.004 mg/ ml). After 30 min at 25uC, the reaction was stopped with 1 ml of ice-cold 25% (w/v) trichloroacetic acid (TCA) and 1 ml of ice-cold 0.05% (w/v) bovine serum albumin. After precipitation on ice for 30 min, the protein precipitate was collected on a 2.5 cm glass filter (Sartorius, Go¨ttingen, Germany) and washed with 10 ml of ice-cold 10% trichloroacetic acid. The amount of 32Pi incorporated in histone was measured by liquid scintillation counting. The linearity of the assay was confirmed from the time-course of histone phosphorylation over a 30 min period. Additional control experiments were run in the absence of calcium to measure basal
kinase activity only adding EGTA without any CaCl2 with a reaction time of 30 minutes.
Data Analysis
The dependence of PKCa activity on the contents of the different activators in the model membranes was analyzed by a non-linear least squares fit to a modified Hill equation:
y~a Vmax
xn K0:5nzxn
where y is the measured activity of PKCa, a is the activity in the absence of lipid or Ca2 (background), Vmax is the lipid-stimulated activity, x is the concentration of the activator, K0.5 is the concentration of activator resulting in half maximal activity and n is the Hill coefficient. Standard errors for n, Vmax and K0.5, taken for three independent experiments, are reported.
Results
The important contribution of PIP2 to PKCa enzymatic activity was clearly observed when it was studied as a function of Ca2+ concentration. A POPC/POPS molar ratio of about 4 was used in these assays since the concentration of POPS in the inner monolayer of eukaryotic plasma membranes, such as in erythrocyte or platelet cells, is roughly this [19–21]. The physiological concentration of PIP2 has been described to be around 1 mol% of the total lipid of plasma membranes [22,23] and it is likely to be concentrated in the inner monolayer at 2 mol%, which increase locally if it forms clusters or patches [24]. As regards diacylglycerol, the physiological levels of this lipid in biomembranes were reviewed in [25]. For example, quantitative measurements of diacylglycerols present in stimulated cells have shown that they may reach 1.45 mol% of the total lipid concentration [26] or about 2 mol% [27]. So the concentrations of diacylglycerol used in this work can be considered physiological and well within the range of diacylglycerol concentrations used in standard procedures for PKC activation assays, which use values similar to those used here [28] or even as high as 11.5 mol% with respect to total lipid [29] or as 19 mol% [30] or 25 mol% [31]. In enzymatic studies where the effect of lipid concentrations were studied, 200 mM Ca2+ was used in order to ensure that this cation was not a limiting factor.
When the dependence of PKCa activity on Ca2+ concentration was studied (Fig. 2) the sigmoidal curves obtained in all cases, pointed to cooperativity. In the presence of POPC/POPS (80:20 molar ratio) alone, increasing concentrations of Ca2+ led to a cooperative increase in activity, with a K0.5 of 1.30 mM in Ca2+ (see Table 1), rising from 107.6 nmol Pi/min/mg at 0.1 mM in Ca2+ to a Vmax of 898.4 nmol Pi/min/mg and a Hill coefficient of 2.28. If DOG was added to the membrane to give a composition of POPC/POPS/DOG (78:20:2 molar ratio) the cooperative behavior was again present, but now the K0.5 was 0.84 mM in Ca2+, with the activity raising from 70.5 nmol Pi/min/mg at 0.1 mM Ca2+ to a Vmax of 1192.7 nmol Pi/min/mg and a Hill coefficient of 2.42. It is clear that in the presence of DOG, the K0.5 for Ca2+ decreases, and there is an increase in Vmax.
When PIP2 was added to the membrane to give a composition of POPC/POPS/PIP2 (75:20:5 molar ratio), K0.5 was now 0.59 mM of Ca2+ (Table 1), the activity was 597.4 nmol Pi/min/
mg at 0.1 mM of Ca2+ and Vmax was 1790.7 nmol Pi/min/mg at 10 mM of Ca2+, the higher cooperativity giving a Hill coefficient of 4.10. Even higher cooperativity (Hill coefficient of 8.61) was observed for a membrane which incorporated also DOG, namely
PC/PS/PIP2/DOG (73:20:5:2 molar ratio), although the activity levels did not change with respect to the membrane without DOG,
being now 512.9 nmol Pi/min/mg at 0.1 mM of Ca2+ while Vmax was 1804.9 nmol Pi/min/mg at 10 mM of Ca2+. It is interesting that at very low concentration of Ca2+, e.g. 0.1 mM, the activity in the presence of PIP2 was higher than in the absence of this phosphoinositide, both in the presence and in the absence of DOG. However, the addition of 2% DOG did not increase the activity levels when 5 mol% PIP2 was present.
The effect of increasing POPS concentration was also studied (Fig. 3). When PIP2 was absent in a POPC/POPS/DOG mixture (98-x:x:2 molar ratio), the effect observed was of positive cooperativity (very high Hill coefficient of 13.18), with a K0.5 =15.12 mol% of POPS (Table 2), which is similar to the effect observed previously for membrane activation of this enzyme [14]. Vmax was 1260.2 nmol Pi/min/mg.
If PIP2 was also present, at just 1 mol%, in a mixture containing POPC/POPS/PIP2/DOG (97-x:x:1:2 molar ratio),positive cooperativity was still observed, with a Hill coefficient of 3.71, an initial activity of 490 nmol Pi/min/mg in the absence of POPS and a Vmax of 1586.2 nmol Pi/min/mg. K0.5 was 15.4 mol% of POPS.
Table 1. Dependence of PKCa activation on Ca2 .
Vmax(nmol Pi/ min/mg) n
Lipid mixture K0.5
POPC:POPS (80:20) 1.3060.08 898.2627.8 2.2860.89 POPC:POPS:DOG (78:20:2) 0.8460.27 1192.7696.3 2.4260.69 POPC:POPS:PIP2 (75:20:5) 0.5860.09 1790.7664.4 4.1060.92 POPC:POPS:PIP2:DOG (73:20:5:2) 0.2660.01 1804.96104.4 8.6160.27 K0.5, n and Vmax were obtained by nonlinear least square fit of the data in the
equation described in the Methods Section. doi:10.1371/journal.pone.0069041.t001
POPS/PIP2/DOG (74-x:25:1:x molar ratio) mixture, K0.5 was
- 0.38 mol% of DOG, which was notably lower than the
- 1.10 mol% observed in the absence of PIP2, while the Hill coefficient showed little change (1.39). When the PIP2 concentration was raised to 5 mol%, to give a POPC/POPS/PIP2/DOG mixture (70-x:25:5:x molar ratio), K0.5 decreased to 0.17 mol% DOG, although Vmax maintained a similar value of 1701.9 nmol Pi/min/mg and the Hill coefficient (0.47) indicated an apparent negative cooperativity.
In another set of experiments, the concentration of PIP2 was varied in the presence and in the absence of DOG, keeping the Ca2+ concentration constant at 200 mM. Fig. 5 depicts the results obtained when the molar percentage in the membrane of PIP2 was increased in the absence of DOG, POPC/POPS/DOG (75x:25:x). As can be seen, the K0.5 was 0.39 (Table 4) and the Vmax 1816.2 nmol Pi/min/mg, with low positive cooperativity (n=1.60). In the presence of 2 mol% with a POPC/POPS/ PIP2/DOG membrane (73-x:25:x:2, molar ratio) K0.5 decreased to 0.11 mol% PIP2. This is interesting since it clearly demonstrates that very low concentrations (well below physiological concentrations) are sufficient to significantly enhance the activity of PKCa.
If PIP2 was present in the lipid mixture, but with no DOG, POPC/POPS/PIP2 (95-x:x:5), the initial activity, even in the absence of POPS, was already high, with a value of 1492 nmol Pi/
min/mg. Vmax reached a value of 2084.1 nmol Pi/min/mg and K0.5 was 13.94 mol% of POPS. Thus the addition of PIP2 decreased K0.5 even if DOG was not present, and the activity was almost saturated and no apparent cooperativity was observed (n=1.10).
- Fig. 3 also shows that when PIP2 was increased to 5 mol%, to give a lipid mixture of POPC/POPS/PIP2/DOG (93-x:x:5:2 molar ratio), a very small increase in activity was already observed when POPS was increased since nearly maximum activity was observed in the absence of POPS (1760 nmol Pi/min/mg) and Vmax was 1895.8 nmol Pi/min/mg, with K0.5 of 8.20 mol% POPS and a Hill coefficient of 1.50.
- Fig. 4 shows the activity studied as function of DOG concentration. When the membrane was composed of POPC/ POPS/DOG (75-x:25:x), K0.5 was 0.82 mol% of DOG (Table 3). The activity was 666 nmol Pi/min/mg at 0 mol% of DOG and rose to give a Vmax of 1307.9 nmol Pi/min/mg and a Hill coefficient of 1.59, indicating low positive cooperativity. When PIP2 was incorporated into this assay at just 1 mol% in a POPC/
Table 2. Dependence of PKCa activation on POPS.
Lipid mixture K0.5 Vmax(nmol Pi/min/mg) n
POPC:POPS:DOG (98-x:x:2) 15.1260.24 1260.2648.4 13.1861.9 POPC:POPS:PIP2:DOG (97-x:x:1:2) 15.4060.71 1586.2671.1 3.7160.77 POPC:POPS:PIP2 (95-x:x:5) 13.9461.45 2084.0673.6 1.1060.39 POPC:POPS:PIP2:DOG (93-x:x:5:2) 8.2060.84 1895.86101.3 1.5060.56 K0.5, n and Vmax were obtained by nonlinear least square fit of the data in the equation described in the Methods Section.
doi:10.1371/journal.pone.0069041.t002
Table 3. Dependence of PKCa activation on DOG.
Lipid mixture K0.5 Vmax(nmol Pi/min/mg) n
POPC:POPS:DOG (75-x:25:x) 0.8260.08 1307.9 52.7 1.5960.22 POPC:POPS:PIP2:DOG (74-x:25:1:x) 0.3860.10 1681.7685.9 1.3960.01 POPC:POPS:PIP2:DOG (70-x:25:5:x) 0.1760.05 1701.9669.2 0.4760.15
K0.5, n and Vmax were obtained by nonlinear least square fit of the data in the equation described in the Methods Section. doi:10.1371/journal.pone.0069041.t003
Vmax was 1857.6 nmol Pi/min/mg in this case, which is not significantly differrent with respect to the mixture without DOG, but the Hill coefficient was now 0.78, indicating apparent negative cooperativity during PIP2 activation.
Discussion
This study was designed to show the importance of PIP2 for the catalytic activity of PKCa. Previous studies have shown that PIP2 binds to a site located in the b3- and b4-sheets of the C2 domain, which is a conserved site found also in other C2 domains (Fig. 1) [8]. It was also shown that the presence of PIP2 considerably increased the kinase activity of PKCa [3]. More recently the binding of PKCa to model membranes was studied by monitoring rapid kinetics [12]. It was observed that, in the absence of PIP2, the protein rapid rate of binding was especially dependent on the POPS concentration and a high affinity complex evolved more slowly, which implies specific interactions with POPS and DOG. Both association and dissociation constants were decreased by the presence of PIP2, but the association equilibrium constant was increased, i.e. the species of PKCa bound to the membrane was favored. A model was proposed in which PKCa binds to the membranes via a two-step mechanism consisting of a rapid initial
recruitment of PKCa to the membrane driven by interactions with POPS and/or PIP2, although interactions with DOG are also involved. PKCa carries out a 2-D search on the lipid bilayer to establish specific interactions with its specific ligands. In this way the longer time of residence of the enzyme in the bound state induced by PIP2 could explain its activating effect.
In this work we have assayed in detail the enzymatic activity of PKCa changing the concentrations of the different activators in order to obtain a comprehensive picture of the way in which PIP2 may affect the catalytic action of this enzyme.
With respect to the dependence on Ca2+, it was observed that, as it has long been known [32], the addition of DOG to POPC/ POPS increased to activity, the Vmax going from 898.2 to 1192.7 nmol Pi/min.mg (Table 1), and decreased K0.5 [33], but the Hill coefficient was not significantly altered, showing positive cooperativity. Therefore, the binding of DOG to the C1 domain will influence Ca2+ binding to the C2 domain, facilitating the binding of the C2 domain to the membrane at lower Ca2+ concentrations, an increase in activity that can only be due to its capacity to retain the C1 domain bound to the membrane, impeding return to the inactive structure of PKC.
The addition of PIP2 in the absence of DOG increased Vmax with respect to a membrane containing POPC/POPS/DOG (1790.7 compared to 1192.7 nmol Pi/min.mg) and the further addition of DOG led to the maximum activation (Vmax reaching 1804.9 nmol Pi/min.mg). However, the effect of DOG in this case was not great since the activity was already close to the maximum in the absence of DOG. Note that when PIP2 is introduced and POPC/POPS/DOG and POPC/POPS/DOG/PIP2 are compared, the K0.5 for Ca2+ is reduced in the presence of PIP2 more than three-fold (from 0.84 to 0.26 mM) and the Hill coefficient increases from 2.42 to 8.61. Since it is known that the presence of PIP2 collaborates in the anchoring of the C2 domain to the membrane [3,8,12,34] and that Ca2+ is necessary to allow the binding of the protein to the membrane, it is clear that there is an interplay between these activators. It seems that the presence of PIP2 lowers the amount of Ca2+ required for binding and activity and, at the same time, increases the cooperativity for the binding of Ca2 . Indeed, it has been shown that PIP2 markedly reduces the
Table 4. Dependence of PKCa activation on PIP2.
Vmax (nmol Pi/min/ mg) n
Lipid mixture K0.5
POPC:POPS:PIP2 (75-x:25:x) 0.3960.61 1816.26143.5 1.6060.03 POPC:POPS:PIP2:DOG (73-x:25:x:2) 0.1160.02 1857.6673.9 0.7860.15
K0.5, n and Vmax were obtained by nonlinear least square fit of the data in the equation described in the Methods Section. doi:10.1371/journal.pone.0069041.t004
concentration of Ca2+ required for the binding of isolated C2 domain [4,34]. It is known that up to 3 Ca2+ ions may bind at the calcium binding site of the C2 domain of PKCa [9,35]. A sequential model for classical PKC membrane binding and activation has been proposed [3,5,36], whereby the increase in intracellular Ca2+ produces the binding of Ca1 and Ca2 when the protein is still in the cytosol, leading to the membrane being targeted by the enzyme through the C2 domain. Ca1 is responsible for bridging the protein to the phospholipid molecules (Fig. 1), which are also recognized with the help of Asn189 and Arg216, whereas Ca2 is responsible for keeping Ca1 in its proper location and for inducing a conformational change in PKC, which partially penetrates and docks in the phospholipid bilayer by means of CBR3 (Arg249 and Thr251). Once recruited to the membrane, a third Ca2+ binds, stabilizing the C2 domainmembrane complex. This enables PKC to reside in the membrane for a longer time, allowing the C1 domain to find the diacylglycerol generated upon receptor stimulation [14,33,37–43].
In the case of PKCa dependence on POPS, Vmax, was clearly increased by the addition of 1 mol% of PIP2, and the addition of 5 mol% in the absence of DOG led to very high activities, which did not increase even when DOG was added. This is an interesting result and confirms the great activation capacity of PIP2, and shows that fixing the C2 domain of PKCa to the membrane through the calcium binding site and the PIP2 site decreases the importance of the C1 domain respect to activity. However, the addition of DOG to the membrane containing 5 mol% PIP2 reduced K0.5 from 13.94 to 8.20 mM, showing that binding of the C1 domain may also play a role.
It has been described that POPS binds cooperatively to PKC, with a stoichiometry of 4 [44], $12 [15] or approximately 8 [45] lipid molecules per molecule of protein. A number of authors have observed apparent cooperativity for the activation in mixed micelles with Triton X-100, leading to high Hill coefficients (higher than 8 [46–48] or about 5 [49]) but when the activation was carried out with lipid vesicles, the Hill coefficients were about
- 2.6 [50] or close to 1 [49]. In our case, a high degree of positive
- 3.71, and at 5 mol% PIP2 no cooperativity was evident. Just one POPS molecule is known to bind to the C2 domain of PKCa [8], although more POPS molecules may bind to the C1 domain [51]. It is interesting in this respect that Hill coefficients close to 1 were observed for the binding of the isolated C2 domain to phospholipid vesicles (A. Torrecillas, Ph.D. Thesis, University of Murcia, 2003).
However, it is nowadays recognized that a number of mechanisms may lead to kinetic cooperativity in the absence of true cooperative interactions, and kinetic models have been suggested to explain the apparent cooperativity observed for the binding of lipid to PKC, for example, proposing ligand trapping [52] or effects specific to the interaction with multiple membrane associated ligands [45] have been suggested, the last authors observing that the apparent cooperativity may be abolished in conditions where membrane binding involves a non-phosphatidylserine mechanism, as in the presence of activators such as phorbol esters. This explains why PIP2 reduces the apparent cooperativity so drastically. Therefore, Hill indexes obtained for the binding of proteins, such as PKC to lipids in vesicles or in micelles, may be informative as regards threshold-binding and how this type of binding may be regulated by different ligands.
Diacylglycerol also plays a role, especially in the absence of PIP2, but in the presence of the phosphoinositide its role is reduced. The reason for that may be related to the tighter anchoring of the enzyme as seen by stopped flow experiments using the full-length enzyme [12] and the different orientation of the C2 domain with respect to the membrane, as seen by studying the membrane docking of this domain [53]. These effects occasioned by the interaction with PIP2 may prolong the activated state. The interplay between DOG and PIP2 was also evident when DOG was changed. Even at 1 mol% of PIP2, K0.5 decreased from 1.10 mol% of DOG in its absence to 0.38 in its presence. If PIP2 was 5 mol%, then K0.5 further decreased to 0.17 mol% of DOG, a substantial decrease compared with the total absence of the phosphoinositide. This illustrates that in the presence of PIP2 the enzyme is tightly bound to the membrane and small concentrations of DOG are sufficient to maintain the activity. It is interesting that small apparent positive cooperativity was detected in the absence of PIP2 (n=1.59), which was reduced following the addition of 1 mol% PIP2. This apparent cooperativity in the binding of DOG may not necessarily reflect that the two C1 subdomains bind to DOG when its concentration is sufficiently high, but may be explained by the apparent cooperativity effect described above for POPS, while PIP2 will reduce the apparent cooperativity due to its increasing of the membrane affinity of the protein. It is remarkable, that at 5 mol% of PIP2 the Hill coefficient (0.47) indicated apparent negative cooperativity for diacylglycerol, which might be explained by a change in membrane structure at relatively high DOG concentrations [25].
It is interesting that very low K0.5 values were observed for PIP2 even in the absence of DOG, the value (0.39 mM) being within the physiological range of concentrations. In the presence of DOG a very reduced K0.5 value of 0.11 mM was observed for PIP2, although Vmax increased very slightly as a result of the addition of DOG, confirming that in the presence of PIP2 diacylglycerol is playing a relatively secondary role in the activation of PKCa. Low KD values have been reported for the binding of PIP2 to the isolated C2 domain of PKCa [4,54] with about 1.9 mM for POPC-POPS-PIP2 vesicles, a value which is compatible with our observations for K0.5.
Taken together, the results show that PIP2 increases the Vmax of PKCa and that when its concentration is 5 mol%, the addition of 2 mol% of DOG does not further increase the activity. Moreover, this concentration decreases K0.5 for Ca2+ more than 3-fold, almost 5-fold that of DOG and by a half that of POPS. It is also noteworthy that K0.5 values for PIP2 amounted to only 0.11 mM in the presence of DOG and 0.39 in its absence, therefore well below the maximum physiological concentration for the inner monolayer of a mammalian plasma membrane. As a consequence, PKCa may be expected to operate near its maximum capacity even in the absence of a cell signal producing diacylglycerol. Nevertheless, we have shown that the presence of DOG may also help, since K0.5 for PIP2 notably fell in its presence. On the other hand, since Ca2+ has been shown to be essential for the binding of PIP2 to the C2 domain of PKCa [4,54], this enzyme may be triggered simply by an increase in the cytoplasm concentration of this cation. Since it has been shown that the other classical isoenzymes of PKC are similar to PKCa as regards to the affinity of their C2 domains for PIP2 [4], the above observations may well be extended to them.
In conclusion, the results obtained in this work are compatible with the sequential mechanism previously proposed (3) and further confirmed in vivo (5). Basically, intracytosolic Ca2+ elevations are the trigger to translocate PKCa to the plasma membrane. Once there, two situations can be found: in microdomains enriched only
with phosphatidylserine, the docking of the C2 domain is not enough to liberate the catalytic domain for substrate access, and as seen in the 3D structure recently solved [55], the C1B domain might still keep blocking the catalytic domain. Due to this, the presence of 1,2-diacyl-sn-glycerol in the lipid vesicles by docking at least the C1A domain enables the enzyme to gain its full activation [56]. A second situation can be found when the microdomains are enriched in phosphatidylserine and PIP2 at the plasma membrane. In this case, the C2 domain docks in a different orientation since it has to anchor through two different points, i.e. the CBR (Ca2+/PS) and the lysine rich cluster (PIP2), this might induce a conformational change that unleash the C1 domain from the blocking conformation and enables the catalytic domain to access the
References
- 1. Corbalan-Garcia S, Gomez-Fernandez JC (2006) Protein kinase C regulatory domains: the art of decoding many different signals in membranes. Biochim Biophys Acta 1761: 633–654.
- 2. Nishizuka Y (1995) Protein kinase C and lipid signaling for sustained cellular responses. FASEB J 9: 484–496.
- 3. Corbalan-Garcia S, Garcia-Garcia J, Rodriguez-Alfaro JA, Gomez-Fernandez JC (2003) A new phosphatidylinositol 4,5-bisphosphate-binding site located in the C2 domain of protein kinase Calpha. J Biol Chem 278: 4972–4980.
- 4. Guerrero-Valero M, Marin-Vicente C, Gomez-Fernandez JC, Corbalan-Garcia S (2007) The C2 domains of classical PKCs are specific PtdIns(4,5)P2-sensing domains with different affinities for membrane binding. J Mol Biol 371: 608– 621.
- 5. Marin-Vicente C, Nicolas FE, Gomez-Fernandez JC, Corbalan-Garcia S (2008) The PtdIns(4,5)P2 ligand itself influences the localization of PKCalpha in the plasma membrane of intact living cells. J Mol Biol 377: 1038–1052.
- 6. Sanchez-Bautista S, Marin-Vicente C, Gomez-Fernandez JC, Corbalan-Garcia S (2006) The C2 domain of PKC alpha is a Ca2+-dependent PtdIns(4,5)P-2 sensing domain: A new insight into an old pathway. Journal of Molecular Biology 362: 901–914.
- 7. Marin-Vicente C, Gomez-Fernandez JC, Corbalan-Garcia S (2005) The ATPdependent membrane localization of protein kinase Calpha is regulated by Ca2+ influx and phosphatidylinositol 4,5-bisphosphate in differentiated PC12 cells. Mol Biol Cell 16: 2848–2861.
- 8. Guerrero-Valero M, Ferrer-Orta C, Querol-Audi J, Marin-Vicente C, Fita I, et al. (2009) Structural and mechanistic insights into the association of PKCalphaC2 domain to PtdIns(4,5)P2. Proc Natl Acad Sci U S A 106: 6603–6607.
- 9. Ochoa WF, Corbalan-Garcia S, Eritja R, Rodriguez-Alfaro JA, GomezFernandez JC, et al. (2002) Additional binding sites for anionic phospholipids and calcium ions in the crystal structures of complexes of the C2 domain of protein kinase C alpha. Journal of Molecular Biology 320: 277–291.
- 10. Ochoa WF, Torrecillas A, Fita I, Verdaguer N, Corbalan-Garcia S, et al. (2003) Retinoic acid binds to the C2-domain of protein kinase C(alpha). Biochemistry 42: 8774–8779.
- 11. Evans JH, Murray D, Leslie CC, Falke JJ (2006) Specific translocation of protein kinase Calpha to the plasma membrane requires both Ca2+ and PIP2 recognition by its C2 domain. Mol Biol Cell 17: 56–66.
- 12. Perez-Lara A, Egea-Jimenez AL, Ausili A, Corbalan-Garcia S, GomezFernandez JC (2012) The membrane binding kinetics of full-length PKCalpha is determined by membrane lipid composition. Biochim Biophys Acta 1821: 1434–1442.
- 13. Verdaguer N, Corbalan-Garcia S, Ochoa WF, Fita I, Gomez-Fernandez JC
- 14. Conesa-Zamora P, Lopez-Andreo MJ, Gomez-Fernandez JC, Corbalan-Garcia S (2001) Identification of the phosphatidylserine binding site in the C2 domain that is important for PKC alpha activation and in vivo cell localization. Biochemistry 40: 13898–13905.
- 15. Newton AC, Koshland DE Jr (1989) High cooperativity, specificity, and multiplicity in the protein kinase C-lipid interaction. J Biol Chem 264: 14909– 14915.
- 16. Hannun YA, Loomis CR, Bell RM (1985) Activation of Protein Kinase-C by Triton X-100 Mixed Micelles Containing Diacylglycerol and Phosphatidylserine. Journal of Biological Chemistry 260: 39–43.
- 17. Orr JW, Newton AC (1992) Interaction of protein kinase C with phosphatidylserine. 2. Specificity and regulation. Biochemistry 31: 4667–4673.
- 18. Chang D, Hsieh PS, Dawson DC (1988) Calcium: a program in BASIC for calculating the composition of solutions with specified free concentrations of calcium, magnesium and other divalent cations. Comput Biol Med 18: 351–366.
- 19. Verkleij AJ, Zwaal RF, Roelofsen B, Comfurius P, Kastelijn D, et al. (1973) The asymmetric distribution of phospholipids in the human red cell membrane. A combined study using phospholipases and freeze-etch electron microscopy. Biochim Biophys Acta 323: 178–193.
substrate and consequently full activation of the enzyme. Whether the C1 domains can interact with the membrane independently of 1,2-diacyl-sn-glycerol is not known but there are previous reports indicating that the C1 domains can interact unspecifically with negatively charged phospholipids through the Arg and Lys residues located in its surface [57].
Author Contributions
Conceived and designed the experiments: JCGF SCG. Performed the experiments: ALEJ APL. Analyzed the data: ALEJ APL SCG JCGF. Contributed reagents/materials/analysis tools: JCGF SCG ALEJ APL. Wrote the paper: JCGF. Enzyme assays: ALEJ APL.
- 20. Chap HJ, Zwaal RF, van Deenen LL (1977) Action of highly purified phospholipases on blood platelets. Evidence for an asymmetric distribution of phospholipids in the surface membrane. Biochim Biophys Acta 467: 146–164.
- 21. Leventis PA, Grinstein S (2010) The distribution and function of phosphatidylserine in cellular membranes. Annu Rev Biophys 39: 407–427.
- 22. McLaughlin S, Wang J, Gambhir A, Murray D (2002) PIP(2) and proteins: interactions, organization, and information flow. Annu Rev Biophys Biomol Struct 31: 151–175.
- 23. Di Paolo G, De Camilli P (2006) Phosphoinositides in cell regulation and membrane dynamics. Nature 443: 651–657.
- 24. Huang S, Lifshitz L, Patki-Kamath V, Tuft R, Fogarty K, et al. (2004) Phosphatidylinositol-4,5-bisphosphate-rich plasma membrane patches organize active zones of endocytosis and ruffling in cultured adipocytes. Mol Cell Biol 24: 9102–9123.
- 25. Gomez-Fernandez JC, Corbalan-Garcia S (2007) Diacylglycerols, multivalent membrane modulators. Chem Phys Lipids 148: 1–25.
- 26. Preiss J, Loomis CR, Bishop WR, Stein R, Niedel JE, et al. (1986) Quantitative measurement of sn-1,2-diacylglycerols present in platelets, hepatocytes, and rasand sis-transformed normal rat kidney cells. J Biol Chem 261: 8597–8600.
- 27. Takuwa N, Takuwa Y, Rasmussen H (1987) A tumour promoter, 12-Otetradecanoylphorbol 13-acetate, increases cellular 1,2-diacylglycerol content through a mechanism other than phosphoinositide hydrolysis in Swiss-mouse 3T3 fibroblasts. Biochem J 243: 647–653.
- 28. Sanchez-Pinera P, Micol V, Corbalan-Garcia S, Gomez-Fernandez JC (1999) A comparative study of the activation of protein kinase C alpha by different diacylglycerol isomers. Biochem J 337 (Pt 3): 387–395.
- 29. Ogita K, Ono Y, Kikkawa U, Nishizuka Y (1991) Expression, separation, and assay of protein kinase C subspecies. Methods Enzymol 200: 228–234.
- 30. Wooten MW, Vandenplas M, Nel AE (1987) Rapid purification of protein kinase C from rat brain. A novel method employing protamine-agarose affinity column chromatography. Eur J Biochem 164: 461–467.
- 31. Bolen EJ, Sando JJ (1992) Effect of phospholipid unsaturation on protein kinase C activation. Biochemistry 31: 5945–5951.
- 32. Takai Y, Kishimoto A, Iwasa Y, Kawahara Y, Mori T, et al. (1979) Calciumdependent activation of a multifunctional protein kinase by membrane phospholipids. J Biol Chem 254: 3692–3695.
- 33. Medkova M, Cho W (1998) Differential membrane-binding and activation mechanisms of protein kinase C-alpha and -epsilon. Biochemistry 37: 4892– 4900.
- 34. Corbin JA, Evans JH, Landgraf KE, Falke JJ (2007) Mechanism of specific membrane targeting by C2 domains: localized pools of target lipids enhance Ca2+ affinity. Biochemistry 46: 4322–4336.
- 35. Torrecillas A, Corbalan-Garcia S, Gomez-Fernandez JC (2004) An infrared spectroscopic study of the secondary structure of protein kinase C alpha and its thermal denaturation. Biochemistry 43: 2332–2344.
- 36. Corbalan-Garcia S, Gomez-Fernandez JC (2006) Protein kinase C regulatory domains: The art of decoding many different signals in membranes. Biochimica Et Biophysica Acta-Molecular and Cell Biology of Lipids 1761: 633–654.
- 37. Oancea E, Meyer T (1998) Protein kinase C as a molecular machine for decoding calcium and diacylglycerol signals. Cell 95: 307–318.
- 38. Corbalan-Garcia S, Rodriguez-Alfaro JA, Gomez-Fernandez JC (1999) Determination of the calcium-binding sites of the C2 domain of protein kinase C alpha that are critical for its translocation to the plasma membrane. Biochemical Journal 337: 513–521.
- 39. Conesa-Zamora P, Gomez-Fernandez JC, Corbalan-Garcia S (2000) The C2 domain of protein kinase calpha is directly involved in the diacylglyceroldependent binding of the C1 domain to the membrane. Biochim Biophys Acta 1487: 246–254.
- 40. Garcia-Garcia J, Corbalan-Garcia S, Gomez-Fernandez JC (1999) Effect of calcium and phosphatidic acid binding on the C2 domain of PKC alpha as studied by Fourier transform infrared spectroscopy. Biochemistry 38: 9667– 9675.
- 41. Bolsover SR, Gomez-Fernandez JC, Corbalan-Garcia S (2003) Role of the Ca2+/phosphatidylserine binding region of the C2 domain in the translocation of protein kinase Calpha to the plasma membrane. J Biol Chem 278: 10282– 10290.
- 42. Nalefski EA, Newton AC (2003) Use of stopped-flow fluorescence spectroscopy to measure rapid membrane binding by protein kinase C. Methods Mol Biol 233: 115–128.
- 43. Feng X, Becker KP, Stribling SD, Peters KG, Hannun YA (2000) Regulation of receptor-mediated protein kinase C membrane trafficking by autophosphorylation. J Biol Chem 275: 17024–17034.
- 44. Ganong BR, Loomis CR, Hannun YA, Bell RM (1986) Specificity and mechanism of protein kinase C activation by sn-1,2-diacylglycerols. Proc Natl Acad Sci U S A 83: 1184–1188.
- 45. Mosior M, Newton AC (1998) Mechanism of the apparent cooperativity in the interaction of protein kinase C with phosphatidylserine. Biochemistry 37: 17271–17279.
- 46. Hannun YA, Loomis CR, Bell RM (1986) Protein kinase C activation in mixed micelles. Mechanistic implications of phospholipid, diacylglycerol, and calcium interdependencies. J Biol Chem 261: 7184–7190.
- 47. Hannun YA, Bell RM (1988) Aminoacridines, potent inhibitors of protein kinase C. J Biol Chem 263: 5124–5131.
- 48. Hannun YA, Loomis CR, Merrill AH Jr, Bell RM (1986) Sphingosine inhibition of protein kinase C activity and of phorbol dibutyrate binding in vitro and in human platelets. J Biol Chem 261: 12604–12609.
- 49. Bazzi MD, Nelsestuen GL (1987) Role of substrate in determining the phospholipid specificity of protein kinase C activation. Biochemistry 26: 5002– 5008.
- 50. Orellana A, Hidalgo PC, Morales MN, Mezzano D, Bronfman M (1990) Palmitoyl-CoA and the acyl-CoA thioester of the carcinogenic peroxisomeproliferator ciprofibrate potentiate diacylglycerol-activated protein kinase C by decreasing the phosphatidylserine requirement of the enzyme. Eur J Biochem 190: 57–61.
- 51. Bittova L, Stahelin RV, Cho W (2001) Roles of ionic residues of the C1 domain in protein kinase C-alpha activation and the origin of phosphatidylserine specificity. J Biol Chem 276: 4218–4226.
- 52. Sandermann H Jr, Duncan TM (1991) Lipid-dependent membrane enzymes. Kinetic modelling of the activation of protein kinase C by phosphatidylserine. Biochim Biophys Acta 1069: 235–240.
- 53. Ausili A, Corbalan-Garcia S, Gomez-Fernandez JC, Marsh D (2011) Membrane docking of the C2 domain from protein kinase Calpha as seen by polarized ATR-IR. The role of PIP. Biochim Biophys Acta 1808: 684–695.
- 54. Sanchez-Bautista S, Marin-Vicente C, Gomez-Fernandez JC, Corbalan-Garcia S (2006) The C2 domain of PKCalpha is a Ca2+ -dependent PtdIns(4,5)P2 sensing domain: a new insight into an old pathway. J Mol Biol 362: 901–914.
- 55. Leonard TA, Rozycki B, Saidi LF, Hummer G, Hurley JH (2011) Crystal structure and allosteric activation of protein kinase C betaII. Cell 144: 55–66.
- 56. Ananthanarayanan B, Stahelin RV, Digman MA, Cho W (2003) Activation mechanisms of conventional protein kinase C isoforms are determined by the ligand affinity and conformational flexibility of their C1 domains. J Biol Chem 278: 46886–46894.
- 57. Sanchez-Bautista S, Corbalan-Garcia S, Perez-Lara A, Gomez-Fernandez JC
(2009) A comparison of the membrane binding properties of C1B domains of PKCgamma, PKCdelta, and PKCepsilon. Biophys J 96: 3638–3647.
Figures
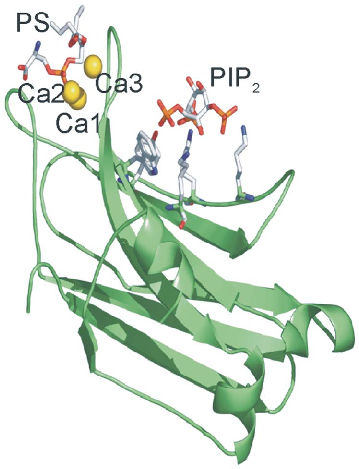
Figure 1
The crystal structure of the PKCα C2 domain bound to calcium, phosphatidylserine, and PIP2 in a quaternary complex is rendered, showing the spatial arrangement of lipid-binding sites.
diagram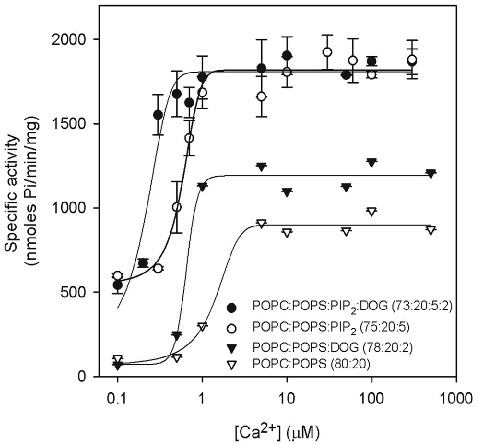
Figure 2
PKCα enzymatic activity is plotted as a function of calcium concentration, demonstrating how PIP2 reduces the calcium requirement for maximal kinase activation.
chart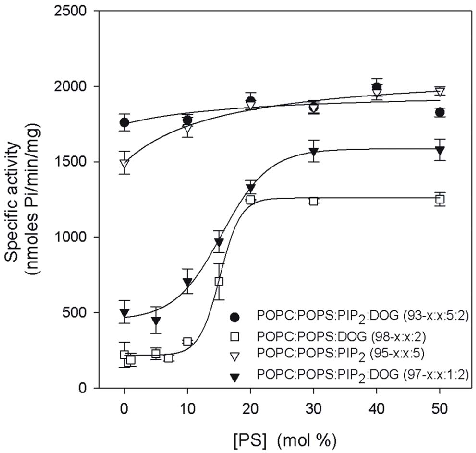
Figure 3
The dependence of PKCα activity on phosphatidylserine (POPS) concentration in lipid vesicles reveals that PIP2 lowers the threshold of phosphatidylserine needed for enzyme activation.
chart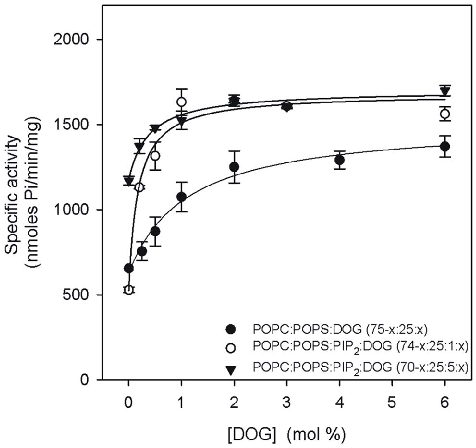
Figure 4
PKCα activity is shown as a function of diacylglycerol (DOG) molar percentage in lipid vesicles, illustrating the cofactor requirements for kinase activation.
chart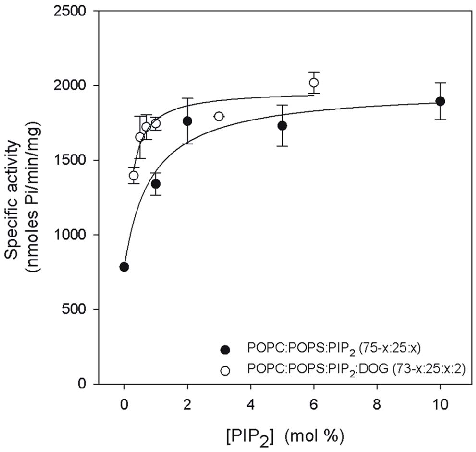
Figure 5
The relationship between PIP2 concentration and PKCα activity demonstrates that increasing PIP2 in lipid vesicles progressively enhances kinase activation efficiency.
chartUsed In Evidence Reviews
Similar Papers
Cellular and molecular life sciences : CMLS · 2005
Surface exposure of phosphatidylserine in pathological cells.
Molecular medicine (Cambridge, Mass.) · 2011
Severe alterations in lipid composition of frontal cortex lipid rafts from Parkinson's disease and incidental Parkinson's disease.
Cell · 1983
Inhibition of VSV binding and infectivity by phosphatidylserine: is phosphatidylserine a VSV-binding site?
Journal of Alzheimer's disease : JAD · 2012
Plasma and brain fatty acid profiles in mild cognitive impairment and Alzheimer's disease.
Current opinion in hematology · 2000
Oxidation and erythrocyte senescence.
FASEB journal : official publication of the Federation of American Societies for Experimental Biology · 2006